According to M. S. Mani (1974), who has studied high altitude environment of most high mountains of the world, high altitude region is the timber line altitude of 2500 to 3000 metres and above. The Arctic region is considered as extending from 60 to 65 degrees north to further northward. The Antarctic region may be taken as extending from 50 degrees south to further southward. But from the present viewpoint the latter polar region is not so important, as at present our knowledge of its insect fauna is very poor.
In the north polar region insects are found mostly up to 79 degrees north. At high altitudes, which are also referred to as the alpine region, insects have been collected up to 4800 metres and above. According to Mani, insects in the alpine zone are mostly concentrated at the snow edge.
The frigid regions are so referred to because of extreme cold in those parts. In the alpine environment the low temperature is due to semirarefied air. Such an atmosphere is very transparent, and its heat retention capacity is low. Presence of suspended particles is very small; it is about 1% of the contents of such particles at the sea level. This further lowers
the capacity of the atmosphere of retaining heat from solar radiation. Air temperature, therefore is very low. In polar regions low atmospheric temperature is not due to rarefied air, but due to the angle of incidence of sun rays. While in the alpine regions the sun rays reach the ground level almost vertically, in polar regions the rays are at a wide deviation from right angles, when approaching the ground.
Another special feature of the alpine region is that the solar radiation is much richer in ultra violet rays (UV) than at the sea level. The dense atmosphere at lower altitudes is quite transparent to the visible part of the solar spectrum, but it is opaque to UV radiation. Hence the radiation reaching the lowland regions is almost free from UV. But in alpine zones UV is in a considerable proportion in the sun light. The same is true for the Antarctic zone due to the ozone hole over the South Pole. While the ozone hole in the south is enlarging through human activity, the ozone layer over the Arctic is thinning out due to a similar reason. Even benign chemicals, released from factories and gadgets, condense on surface of
the polar clouds, and may become ozone destroying.
↚
Insects in frigid regions show a number of interesting structural, physiological, behavioural and developmental adaptations to the extreme conditions in which they live. Both alpine as well as polar region insects tend to be dark due to heavier pigmentation of the integument. They may be black. That is true also in the Andes, where many insects are dark, for instance, some cassidines and some chrysomelines (Elytrosphaera melas in Bolivia and others in Ecuador). They may also be dark blue, dark green or copper. Dark body colours help absorption of heat, when there is sunshine, and they also prevent entry of UV into the body, and thus they protect vital internal organs. In the high tropical mountains, like in the Andes, at the snow level, around 4800 m, there are wingless forms of phasmids, hiding under stones. Many beetles, for instance Metallotimarcha, in European mountains remain active during the night and hide under stones or vegetation during the day. It could be a way to avoid the sun rays. All Metallotimarcha species are copper-like.
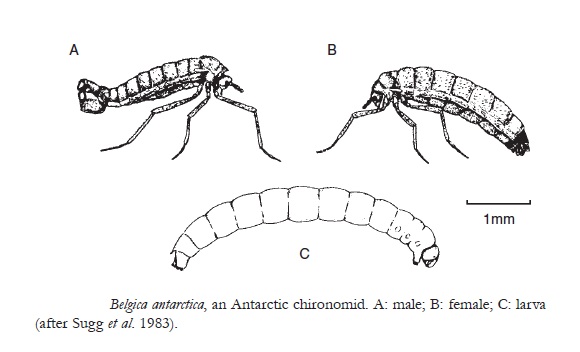
Most alpine and polar species are brachypterous (i.e. with reduced wings) or apterous (i.e. with wings lost). According to Mani, in the north-western Himalayas about 60% of insect species at altitudes above 4000 m are apterous. In the Antarctic Peninsula between 61°S and 65°S, occurs a chironomid, Belgica antarctica (Jolivet, 1991; Sugg et al., 1983), the southernmost free-living holometabolous insect. The adult males and females of this species are apterous. In 1984, a new chironomid species was discovered in the Nepalese Himalayas, a species of Diamesa living on glaciers. The adults of this species have reduced wings and antennae, and are unable to fly. It was found walking on the surface of glaciers (5130 to 5400 m) and in small cavities beneath them (Kohshima, 1984). The larva grows in melt-water drainage channels under the ice and feed on cyanobacteria (Phormidium) and bacteria. The insect spends its entire life cycle in the snow and ice of the glacier, the coldest habitat ever recorded (-16°C). This insect was active at this low temperature. Although several invertebrates have been introduced into the Antarctic, no holometabolous insect has survived there for a long period, except the endemic Belgica. However, a chironomid midge, Eretmoptera murphyi has been recorded from moss at Signy Island, South Orkney Islands (Block et al., 1984). The fly survived for many years and was thought to have been introduced from South Georgia or the Falklands. Its population is parthenogenetic and capable of supercooling to between -13 and -26°C, but it is not active at such low temperatures. Cryoprotectants have been found in the insect extracts.
The loss of wings and flying capacity is a result of natural selection. Alpine and polar regions have violent storms almost regularly; hence selection operates in favour of reduced wings. That windy environs favour reduction of wings has been experimentally shown in Drosophila fly. An entire tropical American genus, Elytrosphaera, linked with high altitudes and with the Brazilian plateau, has fused elytra and is totally wingless. It has normally a bright coloration, but shows darkening with increasing altitude. It is close to the Colorado beetle group, which live in lowlands and are good flyers.
Another advantage of wing loss in beetles (Coleoptera), which dominate among insect fauna of frigid regions, is that, due to disappearance of wings, a subelytral space is created. Such an air-filled space acts as a thermal insulation, and prevents heat loss from the body. (In beetles the front pair of wings have become thick and hard. They are called elytra. The hind wings are membranous, and they alone are used for flight. The elytra form a protective cover for the membranous hind wings in repose. Disappearance of the latter leaves a space beneath the elytra, the subelytral space.) Loss of wings is accompanied by degeneration of flight muscles, a change, which makes room for production of larger eggs. Production of such eggs is a part of the strategy for adaptation to frigid conditions (vide infra).
Still another structural change is reduction in body size. A reduction in surface area of the body reduces heat loss.
Frigid region species remain active when it is sunny. When the sky is overcast and at night, they remain without movements and concealed under grass, weeds and under stones.
A specially notable physiological adaptation in insects in frigid regions is cold resistance. Larvae of the Arctic leaf beetle Chrysolina subsulcata are quite active at -3 to -4 °C. The Himalayan chironomid Diamesa, living on glaciers, is normally active at -16 °C. Springtails (Collembola) are quite numerous around the snow line at high altitudes. They merrily jump about on snow covered fields. Many insects synthetize polyols from glycogen. The cold resistance in these insects is due to presence in their blood of polyols and other anti-freeze substances, similar to those which are mixed with radiator water of cars in cold countries.
Due to paucity of vegetation in frigid areas most insects are debris and carrion feeders. According to Mani on the north-west Himalayas at 5000 metres only 3% species are phytophagous. All the rest are feeders of dead organic matter and are predaceous. Algae and cyanobacteria are the food source for some of them.
Frigid area insects show some interesting developmental adaptations to the cold conditions. Some are viviparous (i.e. the embryonic stages are passed within the body of the mother, and hatched young larvae are given birth to) or ovoviviparous (i.e. early embryonic stages pass within the mother’s body, and eggs with a well developed advanced embryos, ready for hatching, are laid). As a result early embryonic stages are shielded within mother’s body. In general, frigid area insects lay larger eggs (e.g. Brachyhelops, a leaf-beetle in the islands of Southern Patagonia) to permit longer embryonic development due to low temperatures. But Timarcha, which is black, has fused elytra and is totally wingless, lays only a few big eggs and lives mostly in plains. It seems to be a result of a very long evolution, at least from the Jurassic, since the pupae are also wingless. Perhaps Timarcha originated in steppic areas, probably in Central Asia, and it is adapted well to cold in Europe, being black, with a subelytral cavity and having a complex system of diapauses. Quite an interesting exception, and probably a case of preadaptation.
↚
Another developmental adaptation is long periods of hibernation or diapause at more than one developmental stages, so that adverse periods are tided over safely. It may be recalled that food requirement is greatly reduced or is nil during periods of hibernation or diapause. Adult chironomids do not eat generally at the adult stage. As a result of such diapauses the development is a long story. The Arctic species Chrysolina subsulcata shows two larval diapauses. In addition, there may be more diapauses in the larval stage. In this species development from egg to adult stage may take as long as six years (Chernov, 1978; Chernov et al., 1994).
There are no insects in the Antarctic, except two flies and the bird and mammal parasites. Beetles are quite common in the subantarctic islands, but the leaf beetles are missing. There are 21 families of beetles in Greenland, and no chrysomelids, but they were abundant at the Pleistocene (Böcher, 1988). Lack of chrysomelids, in those places, means, except for Antarctic itself, of late no opportunity of dispersion. Leaf beetles are quite capable in surviving in the Southern Greenland climate. In the Antarctic, nematodes, tardigrads and rotifers are quite common living on mosses, lichens and cyanobacteria (Convey and McInnes, 2005). Insects, including beetles, so common during the Jurassic, disappeared with the Nothofagus forests in early Oligocene.
Insects and spiders from plains are often found lying dead at high altitudes. They have been lifted from plains by warm air columns, and have fallen dead and frozen on surfaces high up on mountains. Lowland arthropods may be lifted to high altitudes in considerable numbers. Mani observed in the Himalayas at 4000 m that, in an area of 100 m2, over four hundred dead insects of plains were deposited in 20 minutes during May- June. Bodies of these low land forms should be adding to the food available to carrion feeders at high altitudes.
























